Now Reading: Future of Genomics in Human Forensics
-
01
Future of Genomics in Human Forensics

Recent advances have produced new types of genetic markers with the potential to overcome some important limitations of current DNA profiling methods. These include new molecular approaches for finding individuals previously unknown to investigators, and new molecular methods to support links between forensic sample donors and criminal acts. Such advances in genetics, genomics and molecular biology are likely to improve human forensic case work in the near future1.
DNA profiling with sets of highly polymorphic autosomal short tandem repeat (STR) markers has now been applied in various aspects of human identification in forensic investigations for nearly 20 years2. In brief, autosomal STR profiles are generated from biological materials found at crime scenes and compared with profiles of known suspects identified by police investigations or included in national forensic DNA databases. With statistical support, a profile match provides strong evidence for individual identification, whereas a mismatch does not1.
New tools for identifying known persons
Using autosomal SNPs for human identification
As SNPs reflect single base changes, very short PCR amplicons can be employed, enabling successful SNP profiling from degraded DNA in samples from which STR profiles cannot be obtained3, 4. Furthermore, because SNP variation does not involve repetitive sequences, SNP profiling avoids stutter artefacts that complicate STR profile interpretation, especially when low amounts of DNA are analysed1.
The use of autosomal SNPs for human individual identification in forensic investigations also presents challenges. Obviously, bi‑allelic SNPs are less polymorphic than multi‑allelic STRs. Therefore SNPs are less informative in the analysis of mixtures of DNA from multiple individuals, although the use of a higher number of SNPs relative to STRs3, 4, and/ or perhaps the use of tri‑allelic SNPs5, combined with multiplex genotyping technologies may compensate for this effect1.
Using Y-STRs for male identification
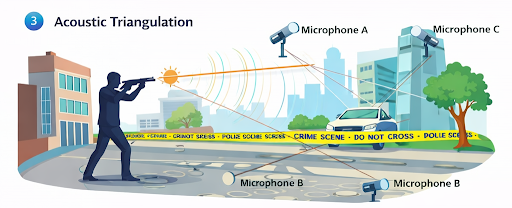
The ability to specifically identify male individuals in cases of sexual assault, where usually mixed stains are analysed that contain DNA from both the male perpetrator and (often in large excess) the female victim, is extremely important6. Haplotypes from sets of non‑recombining male‑specific Y‑STRs have been used for male identification since the late 1990s7, especially once commercial kits containing up to 17 well‑defined Y‑STRs8 became available. Progress has been made in developing frequency databases for these particular Y‑STRs needed for statistical interpretation of Y‑STR profile matches. Y‑STR haplotype databases must be larger than frequency databases for autosomal STRs owing to the non‑recombining nature of the male‑ specific part of the Y chromosome. The largest Y‑STR database is the publically available Y Chromosome Haplotype Reference Database (YHRD)9, which currently includes Y‑STR profiles from nearly 100,000 men from over 700 populations of more than 100 worldwide countries1.
The particular Y‑STR markers currently used in forensics allow the identification of groups of paternally related men (paternal lineages) in most human populations with a high level of resolution6. Exceptions include those populations, such as the Finns10, that have recently undergone bottleneck events or founder events with subsequent expansions. Adding additional Y‑STRs to the current sets can improve the level of paternal line‑ age differentiation, as was recently shown on a worldwide scale11 and also in Finns12. Hence, more markers should be integrated into future commercial Y‑STR kits; suitable Y‑STRs are available, for instance from a comprehensive survey study13.
A general limitation of the particular Y‑STR markers currently applied in forensics is that, unlike autosomal STR profiling, assessment of the weight of evidence is limited for a single contributor as closely related males cannot be excluded. This is because of the low mutation rates of the currently used Y‑STRs, which are in the order of a few mutations per marker every 1,000 generations.14 Such a low mutation rate makes it unlikely that a mutation event occurs between paternally closely related men. However, a recent systematic study that investigated the mutation rate of 186Y‑STRs15 identified 13 markers with much higher mutation rates of a few mutations per marker every 100 generations. With these 13 rapidly mutating (RM) Y‑STRs, it was possible to differentiate >70% of close and distantly related males, whereas the 17Y‑STRs currently used in forensics only differentiated 13% of male relatives from the same samples15. The RM Y‑STRs proved particularly useful for distinguishing between closely related males (separated by 1–5 male generations), for which the currently used Y‑STRs provided almost no differentiation15. RM Y‑STRs are expected to be useful in all cases in which the currently used Y‑STRs did not determine whether a given man or any of his paternal relatives was involved1.
New tools for finding unknown persons
In the absence of a universal DNA profile database, without large enough forensic DNA databases, without efficient policing, and with the ethical, legal and economic problems of DNA mass screenings, one promising approach remains for guiding police investigations to find unknown persons. This approach is known as forensic DNA phenotyping (FDP) and includes the inference of information on biogeographic ancestry and Externally Visible Characteristics (EVCs) directly from a DNA sample. For crime scenes, FDP is expected to reduce the pool of potential suspects and so concentrate and guide police investigations to find the previously unknown perpetrators16.
Inferring biogeographic ancestry from DNA
Large numbers of Y‑SNPs are available for which we have a good understanding of their phylogenetic relationships, a commonly agreed Y haplogroup nomenclature18, and reasonably good knowledge of their global frequency distributions17. These factors make it tempting to use Y‑SNPs for inferring paternal biogeographic ancestry. Global Y chromosome diversity can be captured by genotyping 20–30Y‑SNPs that define the 20 major Y chromosome haplogroups which have differential frequency distributions around the world17, 21. Some of these Y haplogroups are restricted to a single continent: A and B are nearly exclusively found among sub Saharan Africans; H is almost exclusively found on the Indian sub‑continent (and among Roma); and M is almost exclusively found in Oceania. Other Y haplogroups such as R and N are seen across vast areas of Eurasia17. Some geographic knowledge can be obtained from Y‑STRs using global knowledge about haplotype distribution in geo‑referenced databases such as YHRD; however, most Y‑STR haplotypes are too rare to carry geographic information1.
Alternatively, maternal biogeographic ancestry can be inferred from informative mtDNA data. Although the hypervariable region is commonly sequenced for forensic identification purposes, it does not provide enough information for geographic assignment of some mtDNA haplogroups. Recent progress in whole mtDNA sequencing has provided increased understanding about the mtDNA phylogeny and revealed a large number of different mtDNA haplogroups20. Many mtDNA haplogroups show restricted continental distributions, such as haplogroup l to Africa, V to Europe and the Middle East, or P and Q to Oceania17. Genotyping approximately 20–30 coding mtDNA SNPs is likely to allow the definition of most major mtDNA haplogroups with strong frequency differences between geographic regions21.
Estimating appearance traits from DNA
The introduction of genome-wide association studies (GWA studies) for finding genes involved in complex traits provided information for some EVCs such as eye, hair and skin color, pigmentation‑related traits such as freckles22-27, stature, and hair morphology28-30. Furthermore, GWA studies for EVCs involved in disease traits such as nonsyndromic cleft lip with or without cleft palate (NSCl/P)31,32, autosomal‑dominant woolly hair (ADWH)33 and male baldness (androgenetic alopecia (AGA))34,35 may provide genetic leads towards under‑ standing normal appearance variation, although such traits are not directly useful for forensic applications1.
Of all appearance traits studied so far, eye color is the most accurately predictable. A systematic study36 involving thousands of Europeans showed that a model including 15 SNPs predicted eye color with prevalence‑adjusted accuracies of 0.93 for brown and 0.91 for blue eye color as expressed by the area under the receiver characteristic operating curves (AuC), with 1 reflecting complete accuracy and 0.5 random prediction. This is significantly more accurate than an earlier attempt to predict eye color from DNA22, 36. Strikingly, one particular SNP (rs12913832 in HERC2) covers most of the DNA‑predicted eye color information, with AuC values of 0.899 for brown and 0.877 for blue36. From these findings, the IrisPlex system was developed for DNA‑based eye color prediction. This technology includes a highly sensitive multiplex genotyping assay for the six most predictive SNPs from six pigmentation genes, as well as a statistical model with an underlying database that allows reliable categorical eye color prediction independent of biogeographic ancestry37. The IrisPlex assay has already been forensically validated38, and is therefore now available for forensic case work1.
Another trait that is externally visible to some extent and that could be valuable for predicting appearance is individual age. Two DNA‑based approaches for age prediction based on age‑dependant accumulation of mtDNA deletions and age‑dependent telomere shrink‑ age have been suggested for forensic applications, but their practical value seems limited for various reasons39. Genome‑wide studies on age‑dependent changes of gene expression or DNA methylation patterns may pro‑ vide leads for establishing more suitable age‑predictive biomarkers40, 41. The most promising currently known single DNA marker for individual age estimation from blood sources, however, is a particular somatic rearrangement in the T cell receptor locus, particularly its by‑product the signal joint T cell receptor excision circles (sjTRECs)42. Building on previous knowledge that the number of T cells and therefore the number of sjTRECs declines with increasing age, this study showed that normalized sjTREC quantification can be used to estimate age from blood samples with a standard error of ±9 years. It can also be used for categorical age pre‑ diction, with high AuC values from 0.89 to 0.97 for age categories separated by 20 years. The same study described a sensitive quantitative PCR test for sjTREC quantification, which does not seem to be affected by DNA degradation42, although careful forensic validation testing remains to be done1.
Linking sample donors with criminal act
Information that supports a link between sample donors who have been identified by DNA profiling and actual criminal acts is crucial, but until recently attempts to extract such data directly from crime scene samples have been limited1.
Determining cellular origin
Current methods used in forensics for determining the cellular source of a forensic sample, also called forensic tissue identification, are mainly presumptive: that is, they cannot tell us with certainty that a sample is from a particular body source, because positive test results are also possible from other sources43. However, recent developments indicate that this situation might be improved. A study that applied genome‑wide expression microarrays to blood and saliva samples, stored for up to 180 days, revealed sample separation strictly according to body fluids44. Subsequent RT‑PCR confirmation delivered various suitable mRNA markers for the determination of blood and saliva, respectively44. Currently, several mRNA‑ based multiplex RT‑PCR assays are available for the parallel determination of venous blood, saliva, semen and menstrual blood45-48. The value of proposed mRNA markers for vaginal secretion is controversial46,49,50 but may be improved by applying microbial markers51. A recent study identified several mRNA markers and pro‑ vided highly sensitive RT‑PCR methods for skin identification in forensic applications52.
Although some of the mRNA markers proposed for forensic body fluid determination were tested successfully in samples that were up to 16 years old47,53,54, additional environmental conditions such as humidity and temperature are expected to influence mRNA stability, and remain to be tested rigorously1.
As an alternative, microRNA (miRNA) markers have started to be explored for potential use in body fluid determination55, 56. A clear advantage of miRNAs over mRNAs is their small size, 18–22bp, which makes them likely to have higher in vitro stability than mRNAs1.
The advances we have discussed suggest that forensic investigations will improve substantially in the near future. However, several challenges remain. One example is the obvious need for methods to identify multiple donors of a mixed sample, especially when the donors are of the same gender. Here, the use of next‑ generation DNA sequencing technologies from single molecules, especially those not involving PCR, is expected to bring progress. These technologies also provide possibilities for analysing small amounts of degraded DNA, which is important for many forensic cases and for disaster victim identification (DVI)1.
REFERENCES
1. https://www.nist.gov/document/nrg2952pdf
2. Jobling, M. A. & Gill, P. Encoded evidence: DNA in forensic analysis. Nature Rev. Genet. 5, 739–751 (2004).
3. Sanchez, J. J. et al. A multiplex assay with 52 single nucleotide polymorphisms for human identification. Electrophoresis 27, 1713–1724 (2006).
4. Dixon, L. A. et al. Validation of a 21-locus autosomal SNP multiplex for forensic identification purposes. Forensic Sci. Int. 154, 62–77 (2005).
5. Westen, A. A. et al. Tri-allelic SNP markers enable analysis of mixed and degraded DNA samples. Forensic Sci. Int. Genet. 3, 233–241 (2009).
6. Roewer, L. Y chromosome STR typing in crime casework. Forensic Sci. Med. Pathol. 5, 77–84 (2009).
7. Kayser, M. et al. Evaluation of Y-chromosomal STRs: a multicenter study. Int. J. Legal Med. 110, 125–133, 141–149 (1997).
8. Mulero, J. J. et al. Development and validation of the AmpFlSTR Yfiler PCR amplification kit: a male specific, single amplification 17Y-STR multiplex system. J. Forensic Sci. 51, 64–75 (2006).
9. Willuweit, S. & Roewer, L. Y chromosome haplotype reference database (YHRD): update. Forensic Sci. Int. Genet. 1, 83–87 (2007).
10. Hedman, M., Pimenoff, V., Lukka, M., Sistonen, P. & Sajantila, A. Analysis of 16Y STR loci in the Finnish population reveals a local reduction in the diversity of male lineages. Forensic Sci. Int. 142, 37–43 (2004).
11. Vermeulen, M. et al. Improving global and regional resolution of male lineage differentiation by simple single-copy Y-chromosomal short tandem repeat polymorphisms. Forensic Sci. Int. Genet. 3, 205–213 (2009).
12. Hedman, M., Neuvonen, A. M., Sajantila, A. & Palo, J. U. Dissecting the Finnish male uniformity: the value of additional Y-STR loci. Forensic Sci. Int. Genet. 28 Apr 2010 (doi: 10.1016/j.fsigen. 2010.03.007).
13. Kayser, M. et al. A comprehensive survey of human Y-chromosomal microsatellites. Am. J. Hum. Genet. 74, 1183–1197 (2004).
14. Goedbloed, M. et al. Comprehensive mutation analysis of 17Y-chromosomal short tandem repeat polymorphisms included in the AmpFlSTR Yfiler PCR amplification kit. Int. J. Legal Med. 123, 471–482 (2009).
15. Ballantyne, K. N. et al. Mutability of Y-chromosomal microsatellites: rates, characteristics, molecular bases, and forensic implications. Am. J. Hum. Genet. 87, 341–353 (2010).
16. Kayser, M. & Schneider, P. M. DNA-based prediction of human externally visible characteristics in forensics: motivations, scientific challenges, and ethical considerations. Forensic Sci. Int. Genet. 3, 154–161 (2009).
17. Underhill, P. A. & Kivisild, T. Use of Y chromosome and mitochondrial DNA population structure in tracing human migrations. Annu. Rev. Genet. 41, 539–564 (2007)
18. Karafet, T. M. et al. New binary polymorphisms reshape and increase resolution of the human Y chromosomal haplogroup tree. Genome Res. 18, 830–838 (2008).
19. Chiaroni, J., Underhill, P. A. & Cavalli-Sforza, L. L. Y chromosome diversity, human expansion, drift, and cultural evolution. Proc. Natl Acad. Sci. USA 106, 20174–20179 (2009).
20. Van Oven, M. & Kayser, M. Updated comprehensive phylogenetic tree of global human mitochondrial DNA variation. Hum. Mutat. 30, e386–e394 (2009).
21. Behar, D. M. et al. The Genographic Project public participation mitochondrial DNA database. PLoS Genet. 3, e104 (2007).
22. Sulem, P. et al. Genetic determinants of hair, eye and skin pigmentation in Europeans. Nature Genet. 39, 1443–1452 (2007). The first study that investigated human pigmentation with the GWA approach.
23. Kayser, M. et al. Three genome-wide association studies and a linkage analysis identify HERC2 as a human iris color gene. Am. J. Hum. Genet. 82, 411–423 (2008).
24. Han, J. et al. A genome-wide association study identifies novel alleles associated with hair color and skin pigmentation. PLoS Genet. 4, e1000074 (2008).
25. Stokowski, R. P. et al. A genomewide association study of skin pigmentation in a South Asian population. Am. J. Hum. Genet. 81, 1119–1132 (2007).
26. Liu, F. et al. Digital quantification of human eye color highlights genetic association of three new loci. PLoS Genet. 6, e1000934 (2010).
27. Sulem, P. et al. Two newly identified genetic determinants of pigmentation in Europeans. Nature Genet. 40, 835–837 (2008).
28. Medland, S. E. et al. Common variants in the trichohyalin gene are associated with straight hair in Europeans. Am. J. Hum. Genet. 85, 750–755 (2009).
29. Mou, C. et al. Enhanced ectodysplasin-A receptor (EDAR) signaling alters multiple fiber characteristics to produce the East Asian hair form. Hum. Mutat. 29, 1405–1411 (2008).
30. Fujimoto, A. et al. A scan for genetic determinants of human hair morphology: EDAR is associated with Asian hair thickness. Hum. Mol. Genet. 17, 835–843 (2008).
31. Beaty, T. H. et al. A genome-wide association study of cleft lip with and without cleft palate identifies risk variants near MAFB and ABCA4. Nature Genet. 42, 525–529 (2010).
32. Mangold, E. et al. Genome-wide association study identifies two susceptibility loci for nonsyndromic cleft lip with or without cleft palate. Nature Genet. 42, 24–26 (2010).
33. Shimomura, Y., Wajid, M., Petukhova, L., Kurban, M. & Christiano, A. M. Autosomal-dominant woolly hair resulting from disruption of keratin 74 (KRT74), a potential determinant of human hair texture. Am. J. Hum. Genet. 86, 632–638 (2010).
34. Hillmer, A. M. et al. Susceptibility variants for male-pattern baldness on chromosome 20p11. Nature Genet. 40, 1279–1281 (2008).
35. Richards, J. B. et al. Male-pattern baldness susceptibility locus at 20p11. Nature Genet. 40, 1282–1284 (2008).
36. Liu, F. et al. Eye color and the prediction of complex phenotypes from genotypes. Curr. Biol. 19, R192–R193 (2009). A comprehensive study showing that categorical eye colour is predictable with high accuracy from DNA variants.
37. Walsh, S. et al. IrisPlex: a sensitive DNA tool for accurate prediction of blue and brown eye colour in the absence of ancestry information. Forensic Sci. Int. Genet. 27 Mar 2010 (doi:10.1016/j.fsigen. 2010.02.004).
38. Walsh, S. et al. Developmental validation of the IrisPlex system: determination of blue and brown iris colour for forensic intelligence. Forensic Sci. Int. Genet. 12 Oct 2010 (doi:10.1016/j.fsigen. 2010.09.008). This study delivered the first forensically validated DNA test for predicting an externally visible trait, namely eye colour.
39. Meissner, C. & Ritz-Timme, S. Molecular pathology and age estimation. Forensic Sci. Int. 203, 34–43 (2010).
40. Lu, T. et al. Gene regulation and DNA damage in the ageing human brain. Nature 429, 883–891 (2004).
41. Teschendorff, A. E. et al. Age-dependent DNA methylation of genes that are suppressed in stem cells is a hallmark of cancer. Genome Res. 20, 440–446 (2010).
42. Zubakov, D. et al. Estimating human age from T-cell DNA rearrangements. Curr. Biol. 20, R970–R971 (2010). This study showed that categorical age can be accurately estimated from blood, and delivered a DNA test for age estimation.
43. Virkler, K. & Lednev, I. K. Analysis of body fluids for forensic purposes: from laboratory testing to non-destructive rapid confirmatory identification at a crime scene. Forensic Sci. Int. 188, 1–17 (2009).
44. Zubakov, D., Hanekamp, E., Kokshoorn, M., van Ijcken, W. & Kayser, M. Stable RNA markers for identification of blood and saliva stains revealed from whole genome expression analysis of time-wise degraded samples. Int. J. Legal Med. 122, 135–142 (2008).
45. Juusola, J. & Ballantyne, J. mRNA profiling for body fluid identification by multiplex quantitative RT-PCR. J. Forensic Sci. 52, 1252–1262 (2007).
46. Nussbaumer, C., Gharehbaghi-Schnell, E. & Korschineck, I. Messenger RNA profiling: a novel method for body fluid identification by real-time PCR. Forensic Sci. Int. 157, 181–186 (2006).
47. Haas, C., Klesser, B., Maake, C., Bar, W. & Kratzer, A. mRNA profiling for body fluid identification by reverse transcription endpoint PCR and realtime PCR. Forensic Sci. Int. Genet. 3, 80–88 (2009).
48. Fleming, R. I. & Harbison, S. The development of a mRNA multiplex RT-PCR assay for the definitive identification of body fluids. Forensic Sci. Int. Genet. 4, 244–256 (2010). A recent example of the use of mRNA multiplex assays for tissue identification in forensic practice.
49. Juusola, J. & Ballantyne, J. Multiplex mRNA profiling for the identification of body fluids. Forensic Sci. Int. 152, 1–12 (2005).
50. Liu, B. et al. Expression of membrane-associated mucins MUC1 and MUC4 in major human salivary glands. J. Histochem. Cytochem. 50, 811–820 (2002).
51. Fleming, R. I. & Harbison, S. The use of bacteria for the identification of vaginal secretions. Forensic Sci. Int. Genet. 4, 311–315 (2010).
52. Visser, M., Zubakov, D., Ballantyne, K. N. & Kayser, M. mRNA-based skin identification for forensic applications. Int. J. Legal Med. 11 Jan 2011 (doi:10.1007/s00414-010-0545–0542).
53. Zubakov, D., Kokshoorn, M., Kloosterman, A. & Kayser, M. New markers for old stains: stable mRNA markers for blood and saliva identification from up to 16-year-old stains. Int. J. Legal Med. 123, 71–74 (2009).
54. Ferri, G., Bini, C., Ceccardi, S. & Pelotti, S. Successful identification of two years old menstrual bloodstain by using MMP-11 shorter amplicons. J. Forensic Sci. 49, 1387 (2004).
55. Hanson, E. K., Lubenow, H. & Ballantyne, J. Identification of forensically relevant body fluids using a panel of differentially expressed microRNAs. Anal. Biochem. 387, 303–314 (2009). The first study to apply miRNA markers to forensic tissue identification.
56. Zubakov, D. et al. MicroRNA markers for forensic body fluid identification obtained from microarray screening and quantitative RT-PCR confirmation. Int. J. Legal Med. 124, 217–226 (2010).